Arbeitsgruppe Experimentelle Anästhesie
Forschungsschwerpunkte
Die Forschung im Labor der experimentellen Anästhesiologie an der Klinik und Poliklinik für Anästhesiologie widmet sich der Pathophysiologie des Schmerzes, mit dem übergeordneten Ziel molekulare Zielstrukturen zu identifizieren die die Entwicklung von nebenwirkungsarmen und schmerzformspezifischen Arzneimitteln ermöglichen. Dabei werden drei grundlegende Fragestellungen bearbeitet:
1.) Wie werden mechanische Schmerzreize in den Endigungen peripherer Nervenfasern in biochemische bzw. elektrische Signale umgewandelt?
2.) Welche Rolle spielen unterschiedliche Nervenfaseruntergruppen bei der Entstehung unterschiedlicher Schmerzformen (z.B. Entzündungsschmerzen, neuropathische Schmerzen, etc.)?
3.) Wie werden die Schmerzsignale der peripheren Nervenfasern im Rückenmark verarbeitet bzw. zusammengeführt bevor sie an das Gehirn weitergeleitet werden?
Zur Beantwortung dieser Fragen nutzt die Arbeitsgruppe von Prof. Lechner im Tiermodell verschiedenste elektrophysiologische Messtechniken (Patch-Clamp Messungen von Spinalganglien und Rückenmarkschnitten sowie extrazelluläre Aktionspotentialmessungen von einzelnen Nervenfasern) in Kombination mit Optogenetik, bildgebenden Verfahren, molekularbiologischen Techniken, Verhaltenstests, Gentransfer mittels AAV-Viren und transgenen Tiermodellen. Die Forschungsarbeiten werden vorwiegend durch die Deutsche Forschungsgemeinschaft in Form von Sachbeihilfen (LE3210/3-1) und im Rahmen des Sonderforschungsbereich SFB1158 ( www.sfb1158.de ), in dem Prof. Lechner mit zwei Projekten vertreten ist, gefördert.
- Forschung
- Mitarbeiter
- Publikationen
- Partner
-
Forschung
Einzelprojekte
Wie werden mechanische Schmerzreize in den Endigungen peripherer Nervenfasern in biochemische bzw. elektrische Signale umgewandelt?


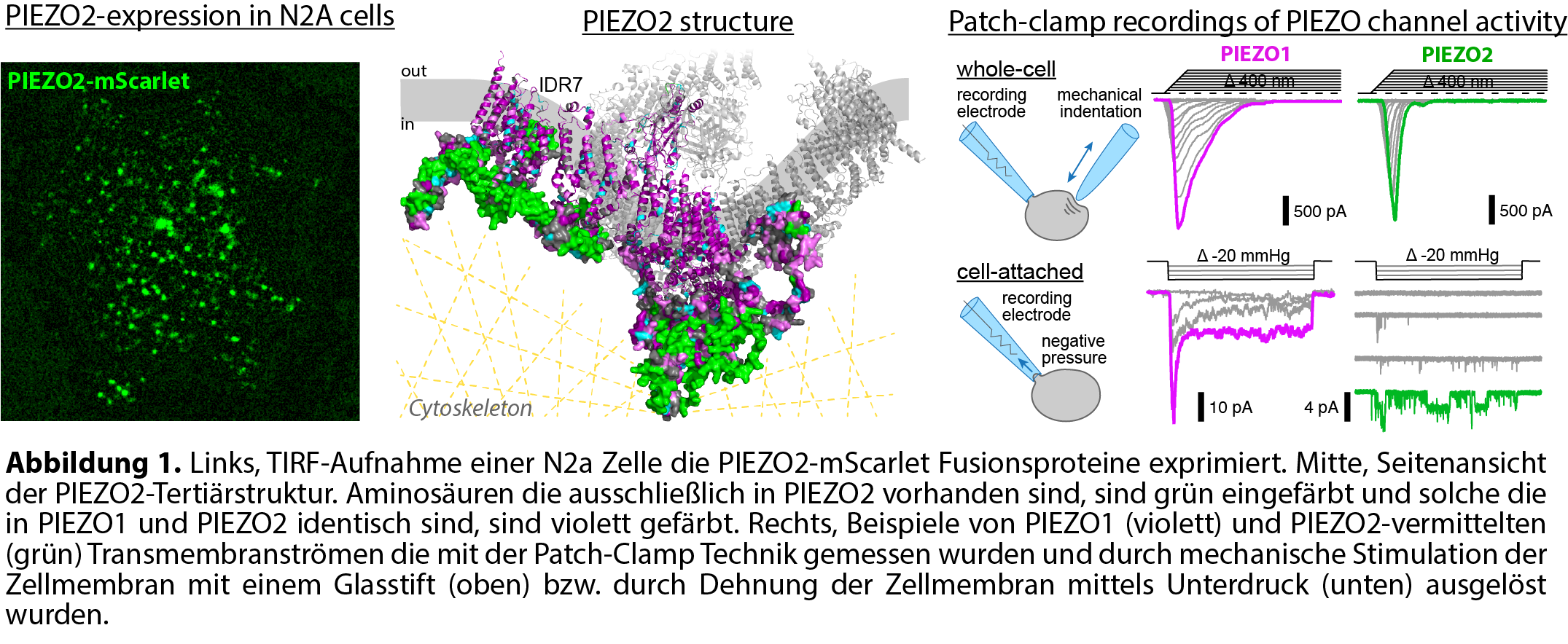
Schmerzhafte mechanische Reize werden von peripheren sensorischen Nervenfasern detektiert und in elektrische Signale umgewandelt – sogenannte Rezeptorpotentiale – welche durch die Nervenfasern an das Rückenmark und von dort an das Gehirn weitergeleitet werden, wo letztlich die Schmerzwahrnehmung erfolgt. Der Umwandlung mechanischer Kräfte in elektrische bzw. biochemische Signale wird als Mechanotransduktion bezeichnet. Die AG Lechner beschäftigt sich seit vielen Jahren mit den molekularen Mechanismen der Mechanotransduktion und konnte in der Vergangenheit unter anderem die embryonale Entwicklung der Mechanotransduktion entschlüsseln [1,2], akzessorische Protein die für die Mechanotransduktion wichtig sind identifizieren [3,4] und zeigen, dass die Umwandlung mechanischer Reize durch Entzündungsmediatoren verstärkt wird, wodurch die Schmerzschwelle sinkt [5]. Derzeit konzentriert sich die AG Lechner mit Blick auf die Mechanotransduktion auf den mechanisch-aktivierbaren Ionenkanal PIEZO2, der nicht nur in sensorischen Nervenzellen [6], sondern auch in einer Vielzahl anderer Zelltypen für die Detektion mechanischer Reize verantwortlich ist. Der Fokus der Arbeiten liegt dabei auf den intrazellulären Kraftübertragungsmechanismen und der Frage welche Proteindomänen der PIEZO-Kanäle für die Kraftübertragung durch das Zytoskelett und die Zellmembran verantwortlich sind [7,8]. Darüber hinaus wird untersucht inwiefern PIEZO-Kanäle die Migration von Zellen bzw. das Längenwachstum von Neuriten und Axonen durch unterschiedlich steife Gewebe kontrollieren. Die Arbeiten zu diesem Thema werden derzeit durch die Deutsche Forschungsgemeinschaft gefördert (LE3210/3-1).
Publikationen der AG Lechner zu diesem Forschungsschwerpunkt:
[1] Lechner et al., EMBO J 28, 1479 (2009).
[2] Wende et al., Science 335, 1373 (2012).
[3] Heidenreich et al., Nature Neuroscience 15, 138 (2012).
[4] Schwaller et al., Nature Neuroscience 24, 74 (2021).
[5] Lechner & Lewin, The Journal of Physiology 587, 3493 (2009).
[6] Schrenk-Siemens et al., Nature Neuroscience 18, 10 (2015).
[7] Taberner et al., PNAS 116, 14260 (2019).
[8] Verkest et al., Nature Communications 13, 1365 (2022).Welche Rolle spielen verschiedene Nervenfaseruntergruppen bei der Entstehung unterschiedlicher Schmerzformen?
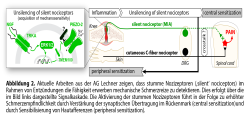
Aus klassischen neurophysiologischen Untersuchungen und jüngeren Transkriptomanalysen ist bekannt, dass periphere sensorische Nerven eine Vielzahl unterschiedlicher Nervenfasern enthalten, die sich in ihren elektrophysiologischen Eigenschaften und ihrem neurochemischen Fingerabdruck unterscheiden. Die AG Lechner untersucht welche Funktion diese verschiedenen Fasertypen bei der Entstehung akuter und chronischer Schmerzen haben. So konnte bereits ein Nervenfasersubtyp identifiziert werden der speziell für die Wahrnehmung von potentiell gewebeschädigenden Reizen, wie z.B. Nadelstichen, verantwortlich ist [1] und gezeigt werden, dass neben Nozizeptoren auch sogenannte Mechanorezeptoren, deren primäre Aufgabe die Detektion von Tastreizen ist, nach Nervenverletzungen zur erhöhten Schmerzempfindlichkeit beitragen [2,3]. Darüber hinaus, hat das Labor Fasertypen beschrieben, die für die für die Regulation der Blutosmolalität und des Blutdrucks wichtig sind [4,5]. Die laufenden Arbeiten im Labor widmen den sogenannten ‚stummen‘ Nozizeptoren, eine Untergruppe von Nozizeptoren, die im gesunden Organismus nicht durch mechanische Reize aktiviert werden, jedoch im Rahmen von Entzündungen in voll funktionsfähige und mechanisch erregbare Nozizeptoren umgewandelt werden. Die AG Lechner konnte vor einigen Jahren einen molekularen Marker für diesen Zelltyp identifizieren [6,7], der die Untersuchung der Rolle der stummen Nozizeptoren bei Entzündungsschmerzen erstmals ermöglichte. Die laufenden Arbeiten des Labors deuten darauf nun hin, dass stumme Nozizeptoren als „Wächter“-Neuronen fungieren die den Zustand des Gewebes überwachen und z.B. im Fall von Entzündungen sekundär adaptive und maladaptive Prozesse, wie Schmerzüberempfindlichkeit in benachbarten Geweben, auslösen [8]. Das Ziel der AG Lechner ist es daher nun, die Aktivierung der stummen Nozizeptoren mittels Gentransfer umzukehren, um die Schmerzausbreitung und somit die Transition von akuten zu chronischen Schmerzen zu verhindern. Die Arbeiten zu diesem Thema werden derzeit durch die Deutsche Forschungsgemeinschaft im Rahmen des SFB1158 Projekt A01 gefördert.
Publikationen der AG Lechner zu diesem Forschungsschwerpunkt:
[1] Arcourt et al., Neuron 93, 179 (2017).
[2] Gangadharan et al., Nature 1 (2022).
[3] Dhandapani et al., Nature Communications 9, 1640 (2018).
[4] Lechner et al., Neuron 69, 332 (2011).
[5] Morelli et al., Cell Reports 35, 109191 (2021).
[6] Prato et al., Cell Reports 21, 3102 (2017).
[7] Schaefer et al., Molecular Pain 14, 1744806918814640 (2018).
[8] Nees et al., bioRxiv, 2022.04.07.486730 (2022).Wie werden die Schmerzsignale der peripheren Nervenfasern im Rückenmark verarbeitet bzw. zusammengeführt bevor sie an das Gehirn weitergeleitet werden?
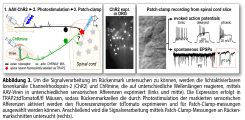
Das Rückenmark spielt eine zentrale Rolle bei der Schmerzentstehung, da hier einerseits die sensorischen Informationen der unterschiedlichen peripheren Nervenfasern zusammengeführt und gefiltert werden, bevor sie an das Gehirn weitergeleitet werden, und andererseits die absteigenden antinozizeptiven Bahnen, die z.B. durch Opioide aktiviert werden, ihre Wirkung entfalten indem sie die zuvor erwähnte Signalverarbeitung modulieren bzw. hemmen. Um die Signalverarbeitung im Rückenmark zu untersuchen, nutzt die AG Lechner Patch-Clamp Messungen von Rückenmarkschnitten in Kombination mit optogenetischen Methoden (siehe Abb. 3). Das übergeordnete Ziel der Arbeiten ist es, die neuronalen Schaltkreise im Rückenmark zu charakterisieren um Zielstrukturen zu identifizieren, die genutzt werden können um die Weiterleitung von Schmerzsignalen und so die Entstehung von Schmerzen zu hemmen. Die Arbeiten zu diesem Thema werden derzeit durch die Deutsche Forschungsgemeinschaft im Rahmen des SFB1158 Projekt A04 gefördert.
-
-
Mitarbeiter
Wissenschaftlicher Leiter·
- Prof. Dr. Stefan Lechner
Wissenschaftliche Mitarbeiter·
- Dr. Clement Verkest
- Dr. Pavel Adamek
- Nadja Zeitzschel MSc
- Lucas Vespermann
Technisches Personal·
- Frau Kirsten Pfeiffer-Drenkhahn
- Frau Claudia Lüchau
- Herr Haider Al-Marsoomi
- Frau Binnur Kuscu
-
Publikationen
Gangadharan, V., Zheng, H., Taberner, F.J., Landry, J., Nees, T.A., Pistolic, J., Agarwal, N., Männich, D., Benes, V., Helmstaedter, M., Ommer B. Lechner S.G., Kuner T. & Kuner R. (2022). Neuropathic pain caused by miswiring and abnormal end organ targeting.NATURE 1–9.
Verkest C., Schaefer I., Nees T.A., Na W., Jegelka J.M., Taberner F.J., & Lechner S.G. (2022). “An intrinsically disordered intracellular domain of PIEZO2 is required for force-from-filament activation of the channel.”NATURE COMMUNICATIONS Mar 15; 13(1) 1365
Morelli C., Castaldi L., Brown S.J., Streich L.L., Websdale A., Taberner F.J., Cerreti B., Barenghi A., Blum K.M., Sawitzke J, Frank T., Steffens L., Doleschall B., Serrao J., Lechner S.G., Prevedel R., Heppenstall P.A. (2021) Identification of a novel population of peripheral sensory neuron that regulates blood pressure.CELL REPORTS Jun 1;35(9):109191
Schwaller, F., Bégay, V., García-García, G., Taberner, F., Moshourab, R., McDonald, B., Docter, T., Kühnemund, J., Ojeda-Alonso, J., Paricio-Montesinos, R., Lechner, S.G., Poulet, J., Millan, J., Lewin, G. (2021). USH2A is a skin end-organ protein necessary for vibration sensing in mice and humansNATURE NEUROSCIENCE 24, 74-81
Agarwal N., Taberner F.J., Rangel Rojas D., Moroni M., Omberbasic D., Njoo C., Andrieux A., Gupta P., Bali K.K., Herpel E., Faghihi F., Fleming T., Dejean A., Lechner S.G., Nawroth P.P., Lewin G.R., Kuner R. (2020) SUMOylation of metabolic enzymes and ion channels in sensory neurons protects against metabolic dysfunction, neuropathy and sensory loss in diabetes.NEURON Sep 23; 107(6):1141-1159
Jonas R., Prato V., Lechner S.G., Groen G., Obreja O., Werland F., Rukwied R., Klusch A., Petersen M., Carr R.W., Schmelz M. (2020) TTX-resistant sodium channels functionally separate silent from polymodal C-nociceptors.FRONTIERS IN CELLULAR NEUROSCIENCE 14(),13
Taberner F.J., Prato, V., Schaefer, I., Schrenk-Siemens, K., Heppenstall P.A. & Lechner, S.G. (2019) Structure-guided examination of the mechanogating mechanisms of PIEZO2.PNAS Jun 24. 116, 14260–14269.
Schäfer I., Prato V., Arcourt A., Taberner F.J. & Lechner S.G. (2018). Differential modulation of voltage-gated sodium channels by nerve growth factor in three major subsets of TrkA-expressing nociceptors.MOL PAIN Nov 2: 1744806918814640
Dhandapani, R., Arokiaraj, C.M., Taberner, F.J., Pacifico, P., Raja, S., Nocchi, L., Portulano, C., Franciosa, F., Maffei, M., Hussain, A.F., Reis, F. de C., Reymond, L., Perlas, E., Garcovich, S., Barth, S., Johnsson, K., Lechner, S.G. & Heppenstall, P.A., (2018). Control of mechanical pain hypersensitivity through ligand-targeted photoablation of TrkB positive sensory neurons.NATURE COMMUNICATIONS 9, 1640
Narayanan P, Huette M, Kudryasheva G, Taberner F.J., Lechner S.G., Rehfeldt F, Gomez-Varela D, Schmidt M. (2018) Myotubularin related protein-2 and its phosphor-lipid substrate PIP2 control Piezo2-mediated mechanotransduction in peripheral sensory neuronseLIFE 7, Mar 9
Prato V.*, Taberner F.J.*, Hockley J.R.F., Callejo G., Arcourt A., Tazir B., Hammer L., Schad P, Heppenstall P.A., Smith E.St.J & Lechner S.G. (2017) Functional and molecular characterization of mechanoinsensitive ‘silent’ nociceptors.CELL REPORTS 21, 3102-3115.,
Arcourt A., Gorham L., Dhandapani R., Prato V., Taberner F.J., Wende H., Gangadharan V., Birchmeier C., Heppenstall P.A. and Lechner S.G. (2017) Touch receptor-derived sensory information alleviates acute mechanical pain and fine-tunes nociceptive reflex coordination.NEURON 93, 179–193.,
Lechner S.G. (2017). An update on the spinal and peripheral pathways of pain signaling.E-NEUROFORUM 23, 131-136,
Lechner S.G. (2017). Shedding light on the contribution of different c-fibre nociceptors to nocifensive behaviour.PAIN Dec; 158(12):2281-2282,
Moshourab, R., Frenzel, H., Lechner, S.G., Haseleu, J., Bégay, V., Omerbašić, D., Lewin, G.R., (2016). Measurement of Vibration Detection Threshold and Tactile Spatial Acuity in Human Subjects.J VIS EXP. Sep 1;(115) ,
Schrenk-Siemens, K., Wende, H., Prato, V., Song, K., Rostock, C., Loewer, A. Utikal, J., Lewin, G.R., Lechner, S.G. and Siemens, J. (2015) PIEZO2 is required for mechanotransduction in human stem cell-derived touch receptors.NATURE NEUROSCIENCE 18, 10-6,
Arcourt, A. and Lechner, S.G. (2015) Peripheral and spinal circuits involved in mechanical allodynia.PAIN Feb 156(2), 220-1,
Lewin G.R., Lechner S.G. Smith E.S. (2014) Nerve Growth Factor and Nociception: From Experimental Embryology to New Analgesic Therapy.HANDBOOK OF EXPERIMENTAL PHARMACOLOGY (220):251-82,
Lechner, S.G., and Lewin, G.R. (2013). Hairy sensation. Physiology (Bethesda) 28, 142–150,
Wende, H., Lechner, S.G., Cheret, C., Bourane, S., Kolanczyk, M.E., Pattyn, A., Reuter, K., Munier, F.L., Carroll, P., Lewin, G.R. and Birchmeier C. (2012). The transcription factor c-Maf controls touch receptor development and function.SCIENCE 335, 1373–1376.,
Heidenreich, M.*, Lechner, S.G.*, Vardanyan, V., Wetzel, C., Cremers, C.W., De Leenheer, E.M., Aránguez, G., Moreno-Pelayo, M.Á., Jentsch, T.J., and Lewin, G.R. (2012). KCNQ4 K(+) channels tune mechanoreceptors for normal touch sensation in mouse and man. (* denotes equal contribution)NATURE NEUROSCIENCE 15, 138–145
Frenzel, H., Bohlender, J., Pinsker, K., Wohlleben, B., Tank, J., Lechner, S.G., Schiska, D., Jaijo, T., Rüschendorf, F., Saar, K., Jordan, J., Millán, J.M., Gross, M. and Lewin G.R. (2012). A Genetic Basis for Mechanosensory Traits in Humans.PLoS BIOLOGY 10, e1001318–754.,
Tappe-Theodor, A., Constantin, C.E., Tegeder, I., Lechner, S.G., Langeslag, M., Lepcynzsky, P., Wirotanseng, R.I., Kurejova, M., Agarwal, N., Nagy, G., Todd, A., Wettschureck, N., Offermanns, S., Kress, M., Lewin, G.R. and Kuner, R. (2012). Gα(q/11) signaling tonically modulates nociceptor function and contributes to activity-dependent sensitization.PAIN 153, 184–196.,
Wende, H., Lechner, S.G., and Birchmeier, C. (2012). The transcription factor c-Maf in sensory neuron development. TRANSCRIPTION Nov 1;3(6) ,
Lechner, S.G., and Siemens, J. (2011). Sensory transduction, the gateway to perception: mechanisms and pathology. EMBO REPORTS. Apr;12(4):292-5,
Lechner, S.G.*, Markworth, S.*, Poole, K., Smith, E.S.J., Lapatsina, L., Frahm, S., May, M., Pischke, S., Suzuki, M., Ibañez-Tallon, I., Luft, F.C., Jordan J. and Lewin G.R. (2011). The molecular and cellular identity of peripheral osmoreceptors. (* denotes equal contribution) NEURON 69, 332–344.
Smith, E.S.J., Omerbašić, D., Lechner, S.G., Anirudhan, G., Lapatsina, L., and Lewin, G.R. (2011). The molecular basis of acid insensitivity in the African naked mole-rat. SCIENCE 334, 1557–1560.,
Poole K., Lechner S.G. and Lewin G.R. The Handbook of Touch: The Molecular and Genetic Basis of Touch. New York: Springer Publications 2011
Lechner, S.G., Frenzel, H., Wang, R., and Lewin, G.R. (2009). Developmental waves of mechanosensitivity acquisition in sensory neuron subtypes during embryonic development.EMBO JOURNAL 28, 1479–1491.,
Lechner, S.G., and Lewin, G.R. (2009). Peripheral sensitisation of nociceptors via G-protein-dependent potentiation of mechanotransduction currents.JOURNAL OF PHYSIOLOGY - LONDON 587, 3493–3503.,
Lechner, S.G., Hussl, S., Schicker, K.W., Drobny, H., and Boehm, S. (2005). Presynaptic inhibition via a phospholipase C- and phosphatidylinositol bisphosphate-dependent regulation of neuronal Ca2+ channels.MOLECULAR PHARMACOLOGY 68, 1387–1396.,
Edelbauer, H., Lechner, S.G., Mayer, M., Scholze, T., and Boehm, S. (2005). Presynaptic inhibition of transmitter release from rat sympathetic neurons by bradykinin.JOURNAL OF NEUROCHEMISTRY 93, 1110–1121.,
Lechner, S.G., Dorostkar, M.M., Mayer, M., Edelbauer, H., Pankevych, H., and Boehm, S. (2004). Autoinhibition of transmitter release from PC12 cells and sympathetic neurons through a P2Y receptor-mediated inhibition of voltage-gated Ca2+ channels.EUROPEAN JOURNAL OF NEUROSCIENCE 20, 2917–28,
Lechner, S.G., and Boehm, S. (2004). Regulation of neuronal ion channels via P2Y receptors.PURINERGIC SIGNALLING 1, 31–41. ,
Lechner, S.G., Mayer, M., and Boehm, S. (2003). Activation of M1 muscarinic receptors triggers transmitter release from rat sympathetic neurons through an inhibition of M-type K+ channels.JOURNAL OF PHYSIOLOGY - LONDON 553, 789–802.,
Kubista, H.*, Lechner, S.G.*, Wolf, A.M., and Boehm, S. (2003). Attenuation of the P2Y receptor-mediated control of neuronal Ca2+ channels in PC12 cells by antithrombotic drugs. (* denotes equal contribution)BRITISH JOURNAL OF PHARMACOLOGY 138, 343–350.
-
Partner
Prof. Dr. Gary R. Lewin, MDC Berlin
Prof. Dr. Martin Schmelz, Universität Heidelberg
Prof. Dr. Marc Freichel, Universität Heidelberg
Prof. Dr. Christian Grimm, LMU München
Prof. Dr. Paul A. Heppenstall, SISSA Triest, Italien